

La selección natural ha diseñado mecanismos biológicos sensibles a diferentes tipos de energía relevante para la supervivencia y la reproducción, son los sistemas sensoriales, también llamado sentidos. Los sentidos son vías por las que la información acerca de parte de la realidad circundante accede y al Sistema Nervioso de las especies que lo poseen. Cualquier tipo de estimulación que pueda influir sobre la conducta, para poder hacerlo, tiene que haber sido transformada en cambios eléctricos neuronales, especialmente en impulsos nerviosos.
1. Introducción a la Fisiología de los Sentidos
Los diferentes tipos de energía susceptibles de influir sobre la conducta de un organismo reciben el nombre genérico de estimulación sensorial.
La vía a través de la cual los estímulos llegan al organismo, esencialmente al SNC y rigen la conducta (o influyen sobre ella) está constituida por los receptores sensoriales localizados en los órganos de los sentidos.
1. Tipos de energía estimular: la modalidad sensorial
La estimulación sensorial constituye el aspecto físico y material de los estímulos, su modalidad (luz, sonido…) y sus características físicas de frecuencia, amplitud o intensidad… No todas las energías afectan al comportamiento ni todas forman parte del acervo de posibles estimulaciones.
Además de detectar y “clasificar” la energía, (extraer la información relativa al tipo de energía), el SN debe ser capaz de codificar su procedencia, intensidad y duración.
A la transformación de las diferentes modalidades energéticas en actividad eléctrica llevada a cabo por los receptores sensoriales se le llama transducción sensorial.
El problema al que se enfrente el SN a la hora de proporcionar información sobre las diferentes modalidades sensoriales es que todas ellas son codificadas en términos eléctricos a nivel neuronal.
Toda actividad eléctrica neuronal, incluido el potencial de acción, es cuantitativa y cualitativamente igual.
¿Cómo es posible que distintas modalidades sensoriales sean codificadas con el mismo código?
La explicación está en que las vías nerviosas que transmiten la información sensorial al SNC llevan esa información a zonas determinadas, diferentes para cada modalidad.
Este planteamiento se conoce como ley de líneas marcadas, expresión con la que se quiere hacer referencia a que las vías sensoriales siguen un trayecto predeterminado y genéticamente programado, desde los receptores a las áreas de proyección sensorial, donde se “interpreta” o descodifica la información recibida.
La modalidad sensorial depende de la zona del SNC a la que llegan los impulsos nerviosos y no de cuál haya sido la energía estimular. Los receptores sensoriales están especialmente “sintonizados” con la modalidad sensorial o tipo de energía a la que responden normalmente.
Cada receptor está programado para responder a un estrecho margen de valores de un tipo determinado de energía: el estímulo adecuado.
2. La cuantificación de la energía estimular: codificación de la intensidad (o amplitud) del estímulo
Las neuronas sensoriales “informan” de la intensidad de la energía incidente, ello hace posible que podamos afirmar que una sensación es más intensa que otra.
2 son los mecanismos con los que cuentan las neuronas sensoriales:
Código de frecuencia
Consistente en que a medida que aumenta la intensidad de la estimulación, aumenta correlativamente la frecuencia de potenciales de acción que una neurona sensorial transmite.
Una neurona no puede generar más de unos 1.000 potenciales de acción por segundo. Cuando una determinada intensidad de estimulación le haga responder con esa frecuencia de disparo, cualquier aumento de esa intensidad pasará desapercibido para esa neurona.
Hay una estrategia que permite ampliar el rango de intensidades codificables sirviéndose de una característica de las neuronas sensoriales: umbral de respuesta.
Cada neurona empieza a responder sólo cuando la estimulación tiene la intensidad necesaria. Este umbral difiere de unas neuronas a otras y mientras que unas responden a estimulaciones muy débiles (son muy sensibles) otras sólo lo hacen cuando la estimulación es intensa o muy intensa.
Fraccionamiento según el rango o código poblacional
La intensidad se codifica no sólo mediante la frecuencia de impulsos nerviosos, sino también considerando el número de neuronas que disparan potenciales de acción y su umbral de disparo. A medida que aumenta la intensidad de la estimulación, las neuronas con un umbral de disparo más alto empiezan a disparar potenciales de acción.
3. La codificación de la duración del estímulo
Un fenómeno bastante habitual es el de la disminución o incluso desaparición de la sensación aun cuando la estimulación sensorial sigue teniendo la misma intensidad.
Cuando esta reducción se explica por la fisiología de los receptores sensoriales recibe el nombre de adaptación sensorial, mientras que cuando los receptores siguen respondiendo de la misma forma, pero el individuo deja de respondes a la estimulación, se suele hablar de habituación, que es un tipo de aprendizaje no asociativo cuya explicación hay que buscarla en cambios fisiológicos en las sinapsis entre neuronas dentro del SNC, o en procesos más complejos.
Hay unos receptores que son más fidedignos que tros, tanto en lo que se refiere a la codificación de la intensidad de la estimulación como a su duración. El caso más característico es el de los receptores de presión táctil asociados a los corpúsculos de Pacini que son receptores (son neuronas) que sólo responden con potenciales de acción al principio y al final de la estimulación y dejan de hacerlo el resto del tiempo, lo que significa que responden a los cambios en el estímulo. Esta reducción es la adaptación sensorial.
El proceso de adaptación sensorial no sucede de la misma forma en todos los receptores sensoriales, algunos se adaptan muy rápidamente, otros tardan mucho en adaptarse o nos e adaptan en absoluto (como es el caso de algunos receptores del dolor).
Los receptores que se adaptan deprisa reciben el nombre de receptores fásicos y los que tardan mucho o no son adaptables se les denomina receptores tónicos.
4. La localización del estímulo: campos receptivos e inhibición lateral
La ubicación o localización del estímulo se basa en la distribución estructural de los receptores sensoriales, en el modo en que se relacionan los receptores sensoriales con las diferentes zonas centrales de procesamiento (SNC) de la información que recogen.
Esto se explica por la diferente densidad de receptores en unas zonas que en otra.
La actividad fisiológica de una neurona sensorial resulta modificada por la estimulación de los receptores que contactan con ella, es lo que se conoce como campo receptivo.
La interacción entre campos receptivos próximos explica el fenómeno denominado inhibición lateral. Es una observación bien contrastada que percibimos mejor los cambios en la intensidad de la estimulación sensorial o zonas de contraste.
La estimulación de un campo receptivo no sólo excita a la neurona sensoria, sino que inhibe a las que tienen campos receptivos próximos.
Esquema representativo de las conexiones neurales que podrían explicar la inhibición lateral:
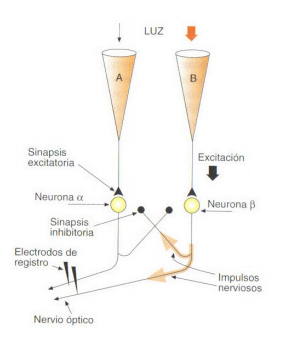
Hay una relación entre campos receptivos, inhibición lateral y frecuencia de impulsos de las neuronas sensoriales: la codificación de la intensidad de un estímulo dado es el resultado de la interacción entre la propia magnitud de ese estímulo y la magnitud de los demás estímulos que puedan estar incidiendo simultáneamente en el campo receptivo de esa neurona.
La intensidad codificada de un estímulo no siempre es directamente proporcional a la intensidad de ese estímulo. La interacción entre campos receptivos también afecta a la frecuencia de disparo de las neuronas sensoriales o de segundo y tercer orden, lo que es una forma de codificar la intensidad del estímulo.
En general los sistemas sensoriales son especialmente efectivos en detectar cambios: la inhibición lateral no es otra cosa que un sistema natural de amplificación.
2. Los Receptores Sensoriales y la Transducción
El estímulo produce cambios en el potencial de membrana de los receptores sensoriales que son de tipo graduado similares a los potenciales postsinàpticos. Este cambio de potencial recibe el nombre de potencial de receptor (o potencial generador) si el receptor sensorial es una neurona.
Este potencial de receptor es una despolarización, aunque los potenciales originados en los receptores visuales son una excepción, pues son hiperpolarizados.
En todas las modalidades sensoriales se han de generar potenciales de receptor que determinaran, bien directa o indirectamente el disparo de potenciales de acción en las neuronas sensoriales.
Para que un organismo pueda verse afectado por una estimulación ambiental, es preciso que el cambio de potencial sobrepase el umbral de disparo de la neurona sensorial y se transforme en un potencial de acción.
A) Quimiorreceptores y Transducción
Receptores gustativos:
Los botones gustativos no se distribuyen de modo uniforme por la lengua, sino que se concentran en las papilas gustativas.
Botón Gustativo:

Las células gustativas proyectan extensiones en forma de microvellosidades hasta el poro en que termina el botón gustativo, mientras que en el otro extremo hacen sinapsis con las neuronas sensoriales.
Existen 4 cualidades gustativas: dulce, salado, amargo y ácido y se añade el glutamato monosódico, el umami.
El sentido del gusto es resultado de la selección natural: las capacidades sensoriales que aporta tienen algún valor adaptativo.
Predecir para qué puede servir el gusto: para distinguir lo que es bueno para comer (y sobrevivir) de lo que no lo es, o sea, qué sustancias exógenas es bueno incorporarlas y qué sustancias serán nocivas para el funcionamiento del organismo.
Transducción gustativa: los receptores gustativos funcionan de forma similar a las neuronas en la medida en que las sustancias estimulantes actúan sobre receptores de membrana, bien abriendo canales iónicos, bien activando receptores proteínicos que inician una cascada metabólica responsable de la activación de segundos mensajeros (según el tipo de sabor, el mecanismo será uno u otro).
En ambos casos el resultado es una despolarización subsecuente a la apertura o cierre de los canales de Na+, K+ o Ca2+ que es de tipo graduado, produciéndose la liberación de un neurotransmisor químico en la sinapsis entre el receptor y la neurona sensorial
Receptores Olfatorios:
Feromonas: son unas sustancias olorosas específicas, capaces de señalar y provocar cambios fisiológicos en los congéneres.
Cavidad nasal con la mucosa olfatoria situada en la parte superior:

Las sustancias olorosas lo son porque algunas de sus moléculas se esparcen por el aire y entran en contacto con los receptores olfativos, neuronas receptoras olfatorias o receptores olfatorios.
En el hombre estos receptores se encuentran en la mucosa olfatoria. Esta mucosa consta de células receptoras (CR), células de soporte (CS) y células basales (CB).
Organización celular de la mucosa olfatoria:

Los receptores olfatorios proyectan cilios (ramificaciones finas de la única dendrita que poseen) hacia el interior de la capa mucosa.
Los receptores olfatorios son neuronas bipolares con una proyección dendrítica hacia la superficie de la mucosa olfatoria donde se engrosa y proyecta sus cilios y una proyección axónica en su polo opuesto que atravesando el hueso craneal a través de la placa cribiforme conecta ipsilateralmente con el bulbo olfatorio.
Los receptores olfatorios cuando interactúan con los estímulos apropiados, responden generando potenciales graduados que se difunden pasivamente por el soma celular hasta alcanzar el segmento proximal del axón, donde se genera un potencial de acción cada vez que se alcanza el umbral.
Se han propuesto 7 olores o aromas básicos o primarios de cuya combinación se derivan todos los demás: etéreo, frutal, floral, canforáceo, menta, pútrido.
La transducción olfativa se inicia en los cilios de las neuronas receptoras, ya que si éstos se eliminan, queda abolida la respuesta olfativa.
La frecuencia de los potenciales de acción depende de la concentración de la sustancia olorosa, pero una exposición prolongada da lugar a una reducción en la frecuencia de los potenciales hasta niveles relativamente bajos, esto se conoce como adaptación sensorial.
Receptores nociceptivos:
Transducción de la información nociceptiva: hay determinados procesos químicos que tienen lugar en los tejidos y afectan a los receptores sensoriales que informan sobre el dolor.
Hay muchos compuestos tanto exógenos como endógenos que afectan a las terminaciones sensoriales nociceptivas. Algunas de estas sustancias activan directamente a las neuronas sensoriales y otras intervienen en los procesos de nocicepción actuando como moduladores modificando el umbral de respuesta de las terminaciones libres a otros estímulos químicos o mecánicos.
Diferentes agentes químicos sensibilizan o estimulan las terminaciones nerviosas libres produciendo dolor. Los tejidos lesionados y las células del sistema inmunitario liberan diversos compuestos que excitan a los nociceptores. Además, la estimulación de los nociceptores puede producir la liberación de sustancias que ellos mismos sintetizan.
B. Mecanorreceptores y transducción
Receptores cutáneos:
Los Mecanorreceptores responden a la acción de fuerzas mecánicas. Entre ellos se encuentran parte de los receptores sensoriales de la piel, y los de los músculos, los tendones y las articulaciones.
Los Corpúsculos de Pacini son receptores que responden a la deformación mecánica de la piel de forma que la presión ejercida por el estímulo genera cambios en la conformación de las láminas del tejido conectivo que rodean la terminación sensorial del receptor, al igual que sucede cuando esta presión se retira.
Estos receptores son neuronas cuya ramificación periférica es la fibra aferente que actúa como receptor sensorial, mientras que su ramificación central responde con potenciales de acción transmitiendo la información al SNC.
Los cuerpos celulares de estas neuronas se encuentran en los ganglios de la raíz dorsal y en los ganglios encefálicos.
Su estructura le hace especialmente sensible para señalar los cambios rápidos de presión.
Corpúsculos de Pacini:

Las sensaciones de la piel se explican por el conjunto de interacciones que se dan a nivel central a partir de la información recogida por los distintos receptores sensoriales. La distribución de los diferentes receptores cutáneos no es homogénea, cada zona tiene una sensibilidad específica, mayor o menos en función de la densidad de receptores.
En las zonas de la piel más sensibles al tacto, como por ejemplo las yemas de los dedos el número de los receptores por área es mayor, un mayor número de neuronas en el SNC que procesan la información procedente de las yemas de los dedos en comparación con otras partes del cuerpo.
La transducción es la transformación de la energía mecánica en potenciales generadores por parte de los corpúsculos de Pacini se lleva a cabo de la siguiente forma:
– el estímulo que incide sobre la piel es modificado por las células accesorias que rodean al receptor
– el estímulo mecánico incide sobre la membrana de la zona receptora de la neurona sensorial para producir un cambio en su potencial de membrana
– este cambio de potencial puede producir un potencial de acción en la fibra nerviosa aferente.
Receptores Auditivos:
La transducción auditiva es el resultado del efecto de las ondas sonoras (fruto de las oscilaciones en la concentración de moléculas del aire que resultan de cualquier proceso vibratorio) que son una forma de fuerza mecánica ejercida por las moléculas del aire, sobre os receptores auditivos: las células ciliadas.
La onda sonora, en su recorrido hasta la cóclea tiene que pasar por el meato auditivo. En su desplazamiento hacia el interior del oído, la onda sonora “tropieza” con la membrana timpánica.
Las variaciones de presión de la onda sonora se transmiten al tímpano que empieza a vibrar con una frecuencia y una amplitud proporcional a la frecuencia e intensidad de la onda sonora que llega.
El tímpano y la cadena de huesecillos del oído medio constituyen una adaptación específica para captar ondas sonoras que viajan a través del aire: originariamente, el oído era una estructura especializada en recoger las vibraciones del medio líquido.
Las vibraciones del aire no son capaces por sí solas de afectar al líquido coclear de los animales terrestres, ya que las ondas sonoras que discurren por el aire apenas afectan a los medios líquidos, sino que se reflejan casi por complejo en ellos.
El tímpano y la cadena de huesecillos que conectan el efecto de la onda o vibración sonora, a través de la ventana oval, con el almacén de líquido que es la cóclea constituyen el sistema de amplificación que permite que las ondas sonoras de tipo aéreo se transmitan a un medio líquido sin apenas pérdida.
El sistema de amplificación se consigue gracias a la interacción de 2 factores:
– la cadena de huesecillos actúa como un sistema de palancas que hace que la vibración del tímpano aumente su fuerza y reduzca su amplitud al llegar a la ventana oval.
– (el más importante) el hecho de que el tímpano tenga una superficie de alrededor de 70mm cuadrados y la ventana oval 3mm cuadrados, lo que determina que la fuerza de la vibración de ésta se vea multiplicada.
Con este sistema se recupera casi por completo la pérdida natural de intensidad por el paso de la vibración de un medio aéreo a un medio líquido.
A. Estructuras esenciales de las 3 partes del oído y B. La representación esquemática del sistema auditivo humano:

Cuando las ondas sonoras llegan a la ventana oval sus vibraciones se transmiten a la perilinfa que rellena la rampa vestibular de la cóclea. Como el líquido coclear no se puede comprimir, cada vez que la ventana oval oscila hacia dentro de la cóclea aumenta la presión del líquido, mientras que cuando oscila hacia fuera, se reduce dicha presión, esto significa que la onda sonora se ha transformado en la cóclea en una onda de presión que recorre la perilinfa líquida.
La vibración de la perilinfa afecta a las 2 membranas que delimitan la rampa media o conducto coclear, relleno de endolinfa.
Estas membranas son:
– la membrana de reissner o vestibular por arriba
– la membrana basilar por abajo.
Ambas membranas sufren una deflección, concretamente la de la membrana basilar.
Visión de un corte de la cóclea en el que se muestran los 2 canales cocleares (rampas vestibulares y timpánica, rellenas de un líquido llamado perilinfa y entre ellas la rampa media o conducto coclear, relleno de endolinfa) así como la deflección que sufren las membranas:

La deflección de la membrana basilar va a ser determinante en el proceso de transducción auditiva, precisamente porque el órgano de Corti, donde se localizan las células ciliadas, se encuentra recubriendo la superficie superior de esta membrana: la incurvación de la membrana basilar por la acción de la vibración de la perilinfa afecta directamente a las células ciliadas. Cuando la membrana basilar se curva, las células ciliadas se curvan paralelamente.
Los estereocilios, que son las vellosidades que poseen las células ciliadas en su parte superior, están en contacto y embebidos en la membrana tectorial, por lo que al incurvarse la membrana basilar, sufren un desplazamiento en su base pero no en la zona de contacto con la membrana tectorial, esto hace que los estereocilios se arqueen más o menos en función de la amplitud de la incurvación de la membrana asilar.
La inclinación de los estereocilios hacia un lado hace que se abran determinados canales iónicos de las células ciliadas de lo que resulta su despolarización.
A. Representación esquemática de la membrana basilar en reposo con el órgano de Corti encima cubierto por la membrana tectorial y B. La membrana basilar se curva como consecuencia de la presión que sobre ella ejerce la perilinfa.

Entre la perilinfa y la endolinfa hay una diferencia de potencial de – 80mV, el llamado potencial endococlear cuyo significado funcional parece ser el de facilitar los cambios de potencial de las células ciliadas. Solo las células ciliadas externas se despolarizan. Las células ciliadas internas tienen muchos menos estereocilios y además no están insertos en la membrana tectorial.
Las células ciliadas hacen sinapsis con las neuronas del nervio auditivo, y las células al despolarizarse liberan cierto neurotransmisor responsable de la despolarización de las neuronas auditivas. Las fibras auditivas están recubiertas de mielina, ésta desaparece en las zonas de contacto con las células ciliadas.
C. Fotorreceptores y Transducción Visual
El procesamiento de la información visual se inicia con la transformación de la luz en señales eléctricas por parte de los fotorreceptores de la retina del ojo.
Los fotorreceptores no producen potenciales de acción tras ser estimulados por la luz, sino que responden con cambios en su potencial de membrana (potenciales locales).
Disposición de los diferentes componentes del ojo humano

Cuando llega la luz la córnea y el cristalino la enfocan y tras atravesar el humor vítreo, es absorbida por las células fotorreceptoras de la retina. Estas células se disponen sobre el epitelio pigmentado que abarca todo el fondo del ojo y contiene grandes cantidades de melanina, cuya función es recoger la luz que las células retinianas no han podido absorber. Ello impide que haya un reflejo de la imagen que entra desde el fondo del ojo hacia la retina, ya que podría distorsionarla.
Para permitir que la luz llegue a las células fotorreceptoras sin ser dispersada o absorbida las capas celulares de la retina más próximas al cristalino no tienen mielina y son relativamente transparentes.
La retina tiene una pequeña invaginación en forma de mella, situada aproximadamente, en su punto central denominada fóvea.
En la fóvea, los cuerpos neuronales de las células retinianas se disponen desplazados hacia los laterales para dejar paso libre a la luz para que no haya distorsión y la luz llegue con menos dificultad.
En la parte central de la fóvea, llamada foveola, prácticamente sólo hay células fotorreceptoras en alta concentración que permite que la agudeza visual sea máxima en esa zona.
Hay 2 tipos de células fotorreceptoras llamadas conos y bastones. Hay más cantidad de bastones que conos. En ambos fotorreceptores existen moléculas sensibles a la luz llamadas pigmentos visuales o fotopigmentos.
Los bastones contienen un tipo de pigmento específico, mientras que hay 3 tipos de pigmentos en los conos que determinan 3 tipos de conos según el pigmento que usan.
Organización de la retina y su disposición en diferentes capas

El pigmento visual de los bastones es más sensible a la luz que los de los conos, puesto que les permite captar más luz, y además, los bastones amplifican más la señal luminosa que los conos. Un único fotón1 puede producir una señal eléctrica detectable en los bastones, mientras que se precisan centenares de fotones para que un cono pueda generar la misma respuesta.
Fotón: es el valor mínimo de la energía luminosa absorbida por la materia. Esta partícula es la portadora de todas las longitudes de onda del espectro electromagnético.
Por lo tanto, los bastones participan en la visión nocturna o con luz tenue, mientras que los conos se activan durante la visión diurna. Tanto los conos como los bastones hacen sinapsis con neuronas bipolares.
Hay muchos bastones que establecen contacto con una misma célula bipolar, reforzando así la señal eléctrica que se genera en ella, pero esta mayor convergencia es una limitación para ganar en resolución espacial porque en la neurona bipolar los efectos de cada bastón se promedian.
Los conos presentan mejor resolución espacial porque muestran menos convergencia (sólo unos pocos establecen sinapsis con cada célula bipolar) y ello facilita que la imagen pueda transmitirse de forma más fidedigna manteniendo las diferencias espaciales. Los conos están concentrados en la fóvea, lugar donde la imagen sufre menor distorsión y donde no hay convergencia de ningún tipo: cada célula bipolar recibe información de un único cono.
También se diferencian en que los conos presentan mayor resolución temporal que los bastones. Los bastones disparan lentamente y permite detectar bajas iluminaciones, en cambio la respuesta de los conos es mucho más rápida, lo que les permite detectar mejor los cambios de la iluminación en el tiempo.
Las diferencias funcionales entre conos y bastones no sólo se deben a las distintas propiedades que presentan ambos fotorreceptores sino también a las conexiones que establecen con otras células de la retina. Estas conexiones constituyen 2 sistemas funcionales distintos en la retina: el escotópico (para los bastones) y el fototópico (para los conos).
Representación de la morfología de las células fotorreceptoras (bastones a la izquierda, conos a la derecha).

Las principales diferencias entre conos y bastones en lo que se refiere a la transducción visual estriban:
– en primer lugar, en su sensibilidad: mientras que los bastones reaccionan a la presencia de un único fotón, los conos requieren mucha mayor cantidad de energía para responder.
Por ello los bastones se saturan y dejan de responder en condiciones de buena iluminación.
– Los conos en cambio son capaces de continuar respondiendo aun cuando se mantenga la luz prolongadamente.
Los mecanismos de la transducción son muy similares en ambos fotorreceptores y que se llevan a cabo de modo parecido a otras formas de transducción basadas en procesos de activación e inactivación de proteínas G y de segundos mensajeros. En la transducción visual el potencial de receptor constituye una excepción a la regla general ya que no se trata de una despolarización sino de una hiperpolarización (importante).
En ausencia de luz, los iones Na+ están entrando masivamente a través de los segmentos externos de los fotorreceptores lo que les mantiene despolarizados, dando como resultado la liberación del neurotransmisor glutamato.
En presencia de luz, los canales de Na+ se cierran y el potencial de membrana se hiperpolariza al no entrar iones Na+, por lo que deja de liberarse masivamente glutamato.
Cuando los fotones de luz son absorbidos por los fotopigmentos, se produce la descomposición de éstos en diversos compuestos a través de una cadena de reacciones moleculares. Uno de estos compuestos activa una proteína G que a su vez activa la degradación del segundo mensajero GMPC.
Este mensajero mantenía abiertos los canales de Na+, lo que mantenía despolarizado al fotorreceptor y al ser degradado el mensajero los canales de Na+ no pueden mantenerse abiertos y los fotorreceptores sufren una hiperpolarización.
3. La transmisión de la información sensorial al sistema nervioso central
El resultado de la transducción es la generación de potenciales de receptor, que finalmente pueden provocar el disparo de potenciales de acción en las neuronas sensoriales, de forma directa o indirecta, según sea el receptor una neurona o una célula especializada no nerviosa. Los axones de estas neuronas sensoriales constituyen los nervios espinales y craneales y a través de ellos la información es transmitida a diferentes divisiones del SNC.
1. Aproximación General a los Circuitos Sensoriales del SNC
Una de las características fundamentales del procesamiento de la información sensorial en el SNC es que se requiere la intervención de diversos núcleos y regiones encefálicas que funcionan de forma integrada. Existe una especialización en las funciones que desempeña cada una de estas regiones encefálicas, de modo que las señales sensoriales fluyen desde niveles inferiores a niveles superiores, estableciéndose una jerarquía en el procesamiento sensorial.
Esta organización jerárquica implica que la información procedente de los receptores sensoriales llega principalmente a la médula espinal y a regiones situadas en el tronco del encéfalo y en el diencéfalo para alcanzar finalmente la corteza cerebral (excepto la información olfativa que llega en primer lugar a los hemisferios cerebrales).
Las áreas corticales constituyen el nivel superior de esta jerarquía y son las responsables del procesamiento más complejo de la información sensorial, así como de su integración e interpretación.
La información relevante sobre los estímulos sensoriales es recogida por receptores especializados situados en los órganos de los sentidos. Esta información es recibida por el SNC a través de vías aferentes que están constituidas por los axones de neuronas sensoriales que en su mayor parte se encuentran agrupadas en ganglios, los ganglios del sistema somático.
Estos ganglios se localizan fuera del SNC y forman parte del SNP, son los ganglios de la raíz dorsal o espinales y los ganglios craneales. Las neuronas que constituyen estos ganglios son las neuronas sensoriales de primer orden o primarias.
Los axones de las neuronas sensoriales primarias (raíces dorsales y fibras aferentes que forman parte de los nervios craneales) establecen sinapsis con neuronas situadas en diferentes divisiones del SNC, que son las neuronas sensoriales de segundo orden o secundarias.
Los axones de estas neuronas secundarias alcanza el tálamo, donde establecen sinapsis con neuronas situadas en diversos núcleos responsables del procesamiento de las señales sensoriales. Estas neuronas talámicas son las neuronas sensoriales de tercer orden y están especializadas según la modalidad sensorial.
Se encuentran formando núcleos concretos en el tálamo que envían las señales de cada modalidad hacia áreas específicas de la corteza cerebral.
Esta organización jerárquica de los circuitos sensoriales implica un procesamiento en serie, que no supone la simple transmisión de información pues en cada relevo tiene lugar una integración sucesiva de las señales sensoriales que es cada vez más compleja a medida que se alcanzan niveles superiores de procesamiento.
Por otro lado, existe también un procesamiento en paralelo de las señales sensoriales, de forma que éstas se transmiten a través de vías múltiples para ser analizadas en circuitos paralelos que parecen desempeñar diferentes funciones.
Ejemplo: el sistema somatosensorial está constituido por 2 circuitos diferentes que transmiten información acerca de distintas características del estímulo:
– el sistema lemniscal: responsable de la transmisión de la información táctil más compleja y precisa, y de las señales propioceptivas.
– el sistema anterolateral: que transmite fundamentalmente las señales nociceptivas y la información sobre la temperatura.
Aunque parecen desempeñar funciones diferentes, en otras hay un cierto grado de solapamiento y ambos sistemas comparten información, por ejemplo, el sistema lemniscal participa en la localización del estímulo doloroso.
Otra de las características de la organización de estas vía sensoriales es que parte de sus fibras aferentes experimentan un cruce hacia el lado contralateral del cuerpo, auque este cruce de fibras o decusación tiene lugar a diferentes niveles según el tipo de señal sensorial.
Ejemplo: en el sistema somatosensorial, 2 de los circuitos que lo integran y que transmiten la información procedente de la médula espinal, el sistema lemniscal y el anterolateral, se decusan a diferentes niveles.
Las fibras aferentes primarias del sistema lemniscal son axones mielinizados de gran tamaño que se disponen ordenadamente para representar con precisión el mapa de la superficie corporal y que ascienden hacia el encéfalo formando las columnas dorsales.
Estos axones ascendentes establecen sinapsis con neuronas situadas en los núcleos de las columnas dorsales del bulbo raquídeo (neuronas secundarias) de ahí que al sistema lemniscal también se le denomine sistema de las columnas dorsales.
Esquema representativo de las 2 vías ascendentes del sistema somatosensorial: el sistema lemniscal y el sistema anterolateral
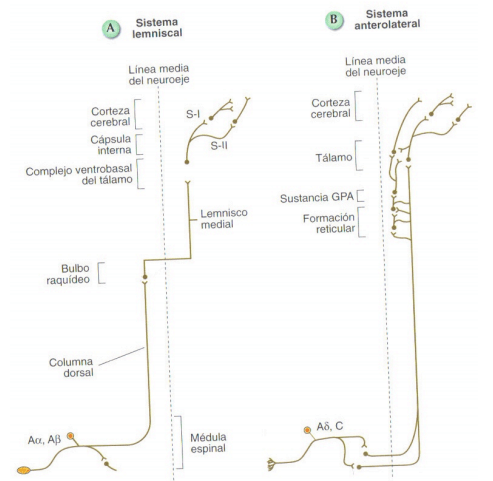
Como puede verse en la figura, los axones de estas neuronas cruzan al lado contrario del cuerpo a nivel bulbar y forman un tracto ascendente denominado lemnisco medial que alcanza en su mayor parte el tálamo y desde esta región diencefálica se envían proyecciones a la corteza somatosensorial.
Las fibras aferentes del sistema anterolateral están constituidas por axones más finos que los del sistema lemniscal, transportan la información más lentamente y permite una menor precisión en la localización del lugar de procedencia de la estimulación.
Estas fibras establecen sinapsis en el asta dorsal de la médula espinal con las neuronas secundarias cuyos axones cruzan al lado opuesto y ascienden a lo largo de la médula espinal (por la zona anterolateral de la sustancia blanca, de ahí su nombre) alcanzando el tálamo y diversas regiones del tronco del encéfalo.
Desde el tálamo la información es enviada, al igual que ocurría con el sistema lemniscal, hasta la corteza somatosensorial y también en este caso la corteza cerebral de cada hemisferio recibe la información procedente del lado contralateral del cuerpo.
Solo en esta vía neural que alcanza la corteza cerebral a través del tálamo se observa el patrón general de organización jerárquica en el que intervienen 3 neuronas (primarias, secundarias, terciarias). El sistema somatosensorial no es el único que presenta una decusación de sus fibras hacia el lado contralateral, otras vías sensoriales, como las auditivas o las vestibulares que decusan a nivel troncoencefálico.
También en el sistema visual encontramos una decusación de las fibras que transmiten la información sobre los estímulos luminosos. Los axones de las células ganglionares de la retina, que constituyen el nervio óptico, se dividen en 2 grupos en el quiasma óptico, de modo que los que provienen de la mitad de la retina más cercana a la nariz (hemirretina nasal) cruzan hacia el lado contralateral, mientras que la otra mitad de la retina (hemirretina temporal) proyecta sus axones ipsilateralmente.
De esta forma la información proveniente de la parte derecha de los campos visuales de ambos ojos se proyecta en la parte izquierda de cada retina, mientras que la información proveniente de la parte izquierda de los campos visuales de ambos ojos se recibe en la mitad derecha de cada retina.
Organización de las vías visuales

A partir del quiasma óptico estos axones constituyen el tracto óptico que transmite la información visual en su mayor parte al núcleo geniculado lateral del tálamo, del que parten axones divergentes que constituyen la radiación óptica que alcanza la corteza visual primaria.
Otra característica de la organización de los sistemas sensoriales de especial relevancia es que presentan una disposición ordenada de sus aferencias que se mantiene en los diferentes niveles de relevo de la información sensorial, desde los niveles inferiores de la jerarquía hasta los niveles superiores.
1. En el sistema somatosensorial, la localización de un estímulo es posible gracias a que la relación existente entre los receptores sensoriales que se encuentran por toda la superficie del cuerpo se mantienen en los niveles superiores de procesamiento en el SNC, es lo que se conoce como organización somatotópica. La organización somatotópica da lugar a un mapa de la superficie corporal que no guarda una relación directa con el tamaño de cada zona, sino que depende de la importancia funcional de esa región.
¿Cómo es posible la organización de este mapa somatotópico? Parte de las fibras aferentes somatosensoriales entran a través de las raíces dorsales a diferentes niveles medulares, de forma que cada raíz dorsal agrupa aferencias procedentes de áreas específicas de la piel denominadas dermatomas.
Dermatoma: es el área de la superficie cutánea inervada por las fibras de cada raíz dorsal.
La organización segmentaria de las raíces dorsales de la médula espinal se mantiene en los diferentes sistemas ascendentes, de forma que las fibras se organizan y terminan en una disposición ordenada que mantiene las relaciones de la superficie corporal según su origen somático.
2. La disposición ordenada que caracteriza la organización de las aferencias sensoriales puede observarse también en el sistema visual. Existe también un mapa retinotópico que permite localizar la situación de los objetos en el campo visual.
Cada punto del campo visual se proyecta en un punto determinado de la retina de forma invertida (lo que está en la parte superior del campo visual se proyecta en la parte inferior de la retina, y lo que está en la parte derecha del campo visual se proyecta en la mitad izquierda de la retina).
La información procedente de cada punto del campo visual es codificada y finalmente transmitida por los axones de las células ganglionares de la retina, que se disponen de forma ordenada para representar el campo visual completo. Esta disposición ordenada de los axones se mantiene también en los niveles superiores del procesamiento visual a nivel talámico y cortical.
La existencia de este mapa retinotópico no implica que se representen de forma proporcionada todas las zonas de la retina (al igual que ocurría con el mapa somatotópico y las zonas corporales) sin que el área de representación es mayor cuanto mayor es la importancia de la región, y éste es el caso de la fóvea o zona central de la retina.
3. También en el sistema auditivo existe una disposición ordenada de los axones aferentes primarios, que nos permite distinguir las diferentes frecuencias de los sonidos. Los axones que establecen contacto con los receptores sensoriales situados en la parte más cercana a la ventana oval codifican sonidos más agudos (frecuencias más altas) mientras que aquellos que establecen sinapsis con los receptores situados más internamente codifican los sonidos más graves (frecuencias más bajas).
Esta organización tonotópica en la que existe una relación topográfica entre los diferentes puntos de procedencia de las fibras auditivas que da lugar a un mapa completo de la cóclea, se mantiene hasta la corteza cerebral.
4. Los Primeros Niveles de Procesamiento de la Información Sensorial
Las fases iniciales del procesamiento de la información visual tienen lugar en la retina. Al conjunto de fotorreceptores que envían información de un área concreta del campo visual a estas células nerviosas se llama campo receptivo de esa célula. El campo receptivo de las células bipolares está constituido por 2 áreas concéntricas, una central en forma de círculo y otra periférica formando un anillo alrededor de la parte central y presenta una organización antagónica.
Representación de los campos receptivos de las células bipolares
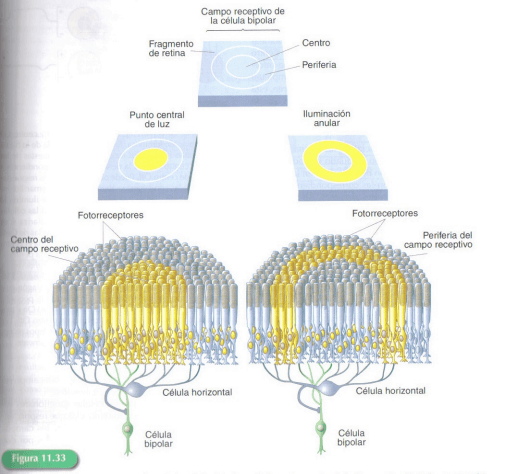
La respuesta de la célula bipolar cuando la luz incide sobre el centro de su campo receptivo es contraria a la que emite cuando la luz incide sobre la periferia.
Se distinguen 2 tipos de células bipolares:
– células de centro ON: si se ilumina el centro del campo receptivo de una célula de centro ON se producirá una despolarización, mientras que la célula se hiperpolarizará si la luz incide en la periferia del campo receptivo.
– células de centro OFF: si se ilumina el centro del campo receptivo de una célula de centro OFF, se producirá una hiperpolarización, pero si la luz incide en la periferia la célula se despolarizará.
Respuesta de las células bipolares de Centro ON y Centro OFF a la estimulación luminosa
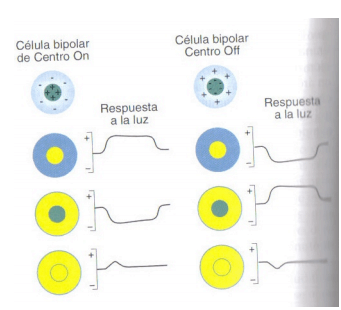
Las células bipolares de Centro ON responden al glutamato liberado por los fotorreceptores hiperpolarizándose, mientras que las de Centro OFF responden al glutamato despolarizándose. En la oscuridad, la liberación de grandes cantidades de glutamato tiene el efecto de hiperpolarizar a las neuronas de Centro ON y despolarizar a las de Centro OFF.
En presencia de la luz, los fotorreceptores liberan menos glutamato, lo que despolariza a la célula de Centro ON y permite la transmisión de información a la célula ganglionar, al mismo tiempo que se produce la hiperpolarización de las células de Centro OFF.
Los terminales de los fotorreceptores pueden establecer contactos sinápticos simultáneamente con células bipolares de Centro ON y de Centro OFF.
Los cambios en el potencial de membrana de las células bipolares son de tipo graduado y producen una mayor o menor liberación de glutamato en las sinapsis que éstas establecen con las correspondientes células ganglionares.
Existen diferentes tipos de células ganglionares según las características del estímulo a las que responden.
Como en el caso de las células bipolares, los campos receptivos de estas células están constituidos por 2 regiones concéntricas con una organización centro-periferia que es antagónica y se nombran como las células bipolares de las que reciben información: células de Centro ON y células de Centro OFF.
Representación de las respuestas de células ganglionares de Centro ON y de Centro OFF.

Estas células responden de forma más vigorosa cuando existe un máximo contraste de la luz que incide en el centro y en la periferia. Cuando sólo se ilumina una parte de su campo receptivo (el centro en las de Centro ON, y la periferia en las de Centro OFF). Las células de Centro ON señalan los aumentos de la iluminación que incide en el centro de su campo receptivo, mientras que las de Centro OFF codifican los decrementos en la iluminación. Por ello se denominan células sensibles al contraste.
La probable función de estas células ganglionares es la de informar al SNC de la diferente distribución de la luz en la retina de las diferencias de contrate en la iluminación, ya que responden de forma débil cuando la iluminación es difusa y afecta a todo su campo receptivo.
5. El papel del tálamo en el procesamiento de la información sensorial
El tálamo es el centro fundamental para el procesamiento de la información sensorial procedente de la médula espinal y del tronco del encéfalo, aunque la información visual llega a él de forma directa. La información olfatoria es una excepción pues alcanza directamente la corteza cerebral, aunque también una parte de ella llega al tálamo.
El tálamo está organizado en diferentes grupos nucleares, entre los que se encuentran los núcleos de relevo sensorial (Figura 7.23 y 7.58), éstos constituyen el grupo ventral y cada un de ellos interviene en el procesamiento de una modalidad sensorial diferente. Estos núcleos envían también de forma específica proyecciones a áreas concretas de la corteza cerebral.
Representación esquemática del tálamo izquierdo para mostrar los núcleos

Figura 7.58: Áreas de la neocorteza (sensoriales, motoras y de asociación) y núcleos talámicos de relevo del grupo ventral que les transmiten las señales

Algunos núcleos talámicos tienen poblaciones celulares muy uniformes, y otros están organizados en capas, como es el caso del sistema visual, las señales llegan al núcleo geniculado lateral, cuyas células se agrupan en función de su tamaño y de las proyecciones que reciben de la retina, en capas parvocelulares (células de pequeño tamaño) y capas magnocelulares (células de gran tamaño) (Figura 11.36)
Figura 11.36 Organización del núcleo geniculado lateral del tálamo

Existen 2 sistemas independientes, aunque también comparten parte de la información, son el sistema parvocelular y el sistema magnocelular. Ambos sistemas proyectan a regiones diferentes de la corteza cerebral. Los campos receptivos de las neuronas talámicas presentan una organización antagónica similar a las de las células bipolares y ganglionares de la retina.
Las neuronas magnocelulares tienen campos receptivos más grandes y son sensibles al movimiento de los objetos y contribuyen a la percepción de las características generales del estímulo, mientras que las neuronas parvocelulares tienen campos receptivos más pequeños y son responsables del análisis de los detalles finos de la imagen e intervienen en la percepción de la forma y del color.
Una vez que las señales visuales han sido procesadas en el tálamo, la información es enviada a la corteza visual a través de las radiaciones ópticas y a otras regiones encefálicas.
6. El procesamiento cortical de la información sensorial
Las proyecciones talámicas llegan a áreas corticales concretas dependiendo de la modalidad sensorial alcanzando, en primer lugar, las áreas sensoriales primarias. Estas áreas transmiten la información a las áreas corticales secundarias y a las áreas de asociación.
1. Procesamiento cortical en el sistema somatosensorial
La corteza somatosensorial esta formada por unas regiones citoarquitectónicamente diferentes situadas en la región anterior del lóbulo parietal.
La corteza somatosensorial primaria se sitúa en la circunvolución post-central y en el interior del surco central, mientras que la corteza somatosensorial secundaria se localiza de forma lateral y algo posterior a la corteza primaria y en el interior del surco lateral.
Figura 11.37 Áreas somatosensoriales primarias y secundarias.

Las áreas somatosensoriales primarias y secundarias de la corteza parietal anterior proyectan a la corteza de asociación parietal posterior. Las lesiones en esta área de asociación no dan lugar a déficit sensoriales específicos como ceguera, sordera o pérdida de la sensibilidad táctil, como ocurriría si se lesionaran las áreas primarias correspondientes.
La organización somatotópica se mantiene también en la corteza somatosensorial. La superficie del cuerpo no está proporcionalmente representada, sino que ésta es mayor para aquellas partes que desempeñan un papel fundamental en la discriminación táctil y de las que hay que tener una información sensorial precisa para poder ejercer un fino control de sus movimientos.
Figura 11.38 Organización somatotópica en la corteza somatosensorial primaria
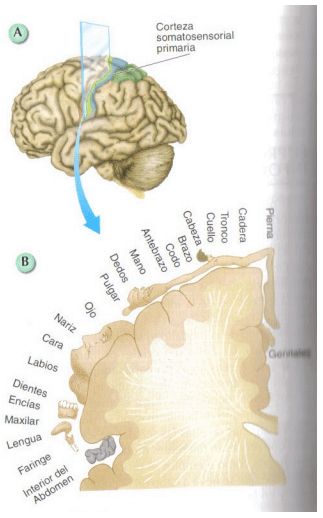
La magnificación cortical de algunas zonas del cuerpo está proporcionalmente relacionada con la densidad de las aferencias sensoriales recibidas desde cada una de ella y éste puede verse modificado en función de la experiencia, (ejemplo: capacidad de discriminación táctil desarrollada en las personas que leen textos en Braille) o por la existencia de lesiones.
La corteza somatosensorial muestra una organización interna, de forma que se establecen conexiones verticales entre las neuronas de regiones concretas, presentando una organización columnar.
Estas columnas están definidas en función de que sus neuronas reciban información de la misma zona y sean sensibles a estímulos similares.
Todas aquellas neuronas corticales que se localizan en una misma columna constituyen un módulo funcional. Dentro de cada columna todas las neuronas responden a la información captada por un mismo tipo de receptor y de la misma zona de la piel. Cada columna cortical codifica una diferente localización y cualidad de la estimulación.
Aunque los diferentes aspectos de la información táctil que llega a la corteza cerebral son procesados en diferentes columnas, en el sistema somatosensorial se produce una integración de la información que incluye todos estos aspectos.
Para integrar los estímulos que nos llegan y relacionarlos con los objetos de nuestro entorno, el SN tiene que combinar la información procedente de muchos Mecanorreceptores que captan diferentes características del estímulo y procesarla en zonas donde convergen diferentes aferencias.
Esta convergencia permite que en niveles sucesivos del procesamiento sensorial las neuronas respondan a características de los estímulos cada vez más complejas y son estas neuronas las que participan en las últimas fases del procesamiento cortical.
Las diferentes áreas de la corteza somatosensorial primaria y secundaria envían proyecciones a la corteza parietal posterior, que es donde se realiza un nivel de procesamiento más elaborado de la información somatosensorial.
Las lesiones en la corteza parietal posterior pueden producir una incapacidad (astereognosia) para reconocer mediante el tacto objetos comunes. Esta corteza parietal posterior integra junto con la información somatosensorial aquella procedente de otros sistemas sensoriales, fundamentalmente del visual.
De esta integración va a depender la percepción de las relaciones espaciales y de la propia imagen corporal. Las lesiones en esta área pueden provocar que el paciente ignore la mitad de su cuerpo, el lado contralateral al hemisferio dañado, el síndrome de heminegligencia.
2. Procesamiento cortical en el sistema visual
La información visual llegar a través de los tractos ópticos al tálamo, concretamente, al núcleo geniculado lateral y desde allí es enviada mediante las radiaciones ópticas a un área concreta de la corteza cerebral situada en el lóbulo occipital, la corteza visual primaria (V1), aunque existen también otras áreas de la corteza visual denominadas V2, V3, V4 y V5 que procesan también diversos aspectos de la información visual.
Figura 11.40 Localización de las diferentes áreas de la corteza visual en el encéfalo. A. distribución general de esas áreas en el lóbulo occipital.

2. A. Las células simples y complejas
Se distinguen 2 tipos principales de neuronas en la corteza visual primaria: las células simples y las células complejas.
Al igual que las células bipolares y ganglionares de la retina y las del núcleo geniculado lateral, las células simples presentan campos receptivos con una zona central y una zona periférica. Los estímulos que son óptimos para la activación de las células simples no son puntos anillos de luz, sino más bien estímulos del tipo de barras alargadas.
Cada célula simple responde a una orientación característica del estímulo en su campo receptivo y que, al mismo tiempo, todas las orientaciones posibles del estímulo están presentes en el conjunto global de la población de las células simples.
No todos los campos receptivos de las células simples presentan una zona central definida como excitadora y una periferia que rodea a esa zona central como inhibidora:
– Hay células simples que tienen campos receptivos con una zona excitadora que ocupa la mayor parte del mismo, mientras que la zona inhibidora ocupa solamente un pequeño lateral.
– Hay células simples que tienen campos receptivos con una zona excitadora ocupando exactamente la mitad del mismo y una zona inhibidora ocupando la otra mitad.
El siguiente nivel del procesamiento de la información visual parece que es llevado a cabo por las células complejas de la corteza visual primaria. Existen diferentes tipos de células complejas que están organizadas jerárquicamente dependiendo de las características de los estímulos que son capaces de desencadenar su activación.
Estas células son las más abundantes de la corteza visual y se localizan no solamente en el área V1, sino también en la V2 y en otras áreas de la corteza visual, a diferencia de las células simples que están agrupadas en su mayor parte en el área V1.
Las células complejas no tienen campos receptivos organizados en áreas excitadoras o inhibidoras ni son activadas por estímulos fijos de luz ya sean puntos o barras. Este tipo de neuronas son estimuladas por el movimiento brusco de un hilo o una barra de luz alargada que recorre su campo receptivo, pero con una orientación determinada.
Las células complejas tienen unos campos receptivos más grandes que los de las células simples, lo que sugiere que estos campos receptivos son el resultado de aferencias provenientes de diversas células simples, que están organizadas espacialmente de una forma característica.
Dado que las células complejas responden de forma intensa ante movimientos del estímulo luminoso en una dirección y no responden en absoluto en otras, se dice que tienen la propiedad de selectividad de la dirección.
Otra propiedad de estas células corticales es la finalización del campo receptivo, ya que la mayoría de estas células responden mucho mejor ante un estímulo cuyo tamaño se ajusta a su campo receptivo, siendo su respuesta más débil cuando la barra de luz es más corta que el propio campo receptivo.
La propiedad de finalización del campo receptivo pueden presentarla también las células simples, pero esta característica es más común en las células complejas del área V2.
El hecho de que los estímulos se muevan parece ser de gran importancia para el procesamiento de la información visual en la corteza cerebral. Si las imágenes fueran perfectamente fijas, al cabo de poco tiempo no se percibirían.
2. B. Las Columnas y los Módulos Corticales
En la corteza visual primaria la mayoría de las neuronas, ya sean simples complejas reciben información procedente de los 2 ojos, son binoculares. Sin embargo, la mayoría de ellas reciben más aferencias desde un ojo que desde el otro, propiedad que se denomina dominancia ocular.
Las neuronas que responden de forma preferente a las señales procedentes de un ojo no se distribuyen al azar en la corteza visual. Estas células se organizan en columna de 1mm de anchura llamadas columnas de dominancia ocular.
Existe también una organización columnar para la preferencia por la orientación de los estímulos con un ángulo determinado ya que las neuronas (simples o complejas) que responden a la misma orientación del estímulo se sitúan en la misma columna, formando las denominadas columnas de orientación.
Figura 11.45 Organización de los módulos corticales visuales.

Al conjunto de las columnas que cubren todos los ángulos posibles de orientación de un estímulo se les denomina hipercolumnas.
Las agrupaciones neuronales o regiones sensibles al color se denominan estacas o manchas y se disponen exactamente alineadas en las columnas de dominancia ocular atravesando las capas de la corteza primaria. Muchas de las células de las estacas son oponentes al color. Las células corticales muestran una organización antagónica centro-periferia y por ello se denominan doblemente oponentes al color.
Las columnas de dominancia ocular y las de orientación se disponen formando ángulos entre sí, de forma que pueden hacerse bloques de corteza que contienen todos los elementos neurales necesarios para el procesamiento de la información visual procedente de la retina. A estos bloques de corteza se les denomina módulos corticales.
Los módulos corticales son las entidades individuales mínimas capaces de procesar la información sobre todos los aspectos de las imágenes que llega a la corteza visual primaria procedente de la retina. Todos los aspectos de los estímulos recibidos en una zona particular de la retina, serían procesados en un módulo concreto. En otro módulo contiguo, se procesaría la información proveniente de otra zona de la retina, y así sucesivamente.
Los módulos corticales son iguales en cuanto a su organización celular, aferencias y eferencias y la forma de analizar la información que reciben. Miles de estos módulos procesarían en conjunto y en paralelo la imagen que se recoge en la retina.
El tipo de células complejas, al requerir un alineamiento exacto del estímulo, participan en la percepción de la profundidad. Por otro lado, dado que los ojos están ligeramente separados, las imágenes que vemos inciden sobre partes ligeramente diferentes de los campos receptivos de las neuronas corticales.
Esta pequeña diferencia en la posición de los objetos es detectada por ciertas células complejas, denominadas células moduladoras de la disparidad.
7. Vías de Modulación Sensorial: La modulación de la percepción del dolor
Las proyecciones descendientes modulan el flujo de información sensorial que alcanza la corteza cerebral, actuando principalmente a nivel del tálamo y del tronco del encéfalo.
La existencia de mecanismos neurales que reducen la sensibilidad al dolor, de modo que la información nociceptiva puede ser modulada tanto por las aferencias sensoriales nociceptivas que recibe la médula espinal como por la activación de vías descendentes procedentes de diferentes estructuras del encéfalo que alcanzan la médula espinal y el tronco del encéfalo.
Las fibras nociceptivas llegan al asta dorsal de la médula espinal donde establecen sinapsis con diferentes tipos de neuronas, parte de cuyos axones constituyen las vías ascendentes que transmiten la información nociceptiva a centros superiores del SNC (sistema anterolateral).
De las vías que constituyen este sistema, el tracto espinotalámico se sabe que su lesión causa un déficit en la sensación de dolor y su estimulación la produce. En la localización del dolor interviene el sistema lemniscal.
No parece existir una organización de las aferencias nociceptivas a la corteza cerebral similar a la de los mapas somatotópicos de las aferencias táctiles. Los núcleos talámicos que reciben información nociceptiva desde las neuronas espinales proyectan difusamente a diferentes regiones de la corteza somatosensorial.
Hay mecanismos en el SN que modulan la transmisión del dolor y modifican la reacción emocional al mismo. Los factores emocionales, cognitivos y socioculturales pueden modificar la percepción del dolor y explicar la variabilidad de respuestas que damos a la estimulación nociceptiva.
En la médula espinal, la actividad de las neuronas que reciben información nociceptiva podía ser modulada por otras aferencias sensoriales que convergen sobre ellas, concretamente, por la estimulación cutánea no nociceptiva, que es capaz de disminuir o bloquear el dolor.
Esto es debido a que la estimulación de las fibras procedentes de los Mecanorreceptores hace disminuir la respuesta de las neuronas del asta doral a la información que llega a través de las fibras nociceptivas.
Figura 11.46 Circuito de modulación de la transmisión de información nociceptiva planteado por la Teoría del control de entrada.
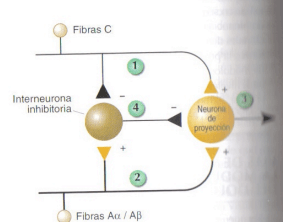
La transmisión de la información sobre el dolor en las neuronas espinales parece no depende exclusivamente de las aferencias nociceptivas, sino de la interacción de éstas con otras fibras aferentes no implicadas directamente en la transmisión de información sobre el dolor. Se ha propuesto la existencia de un circuito neural específico que modula la transmisión de la información nociceptiva que se conoce como Teoría del control de entrada del dolor.
Desde diversos núcleos del encéfalo parten vías descendentes hasta la médula espinal que modulan la transmisión de la información sobre el dolor
Figura 11.47 Vías descendentes de modulación de la información nociceptiva.
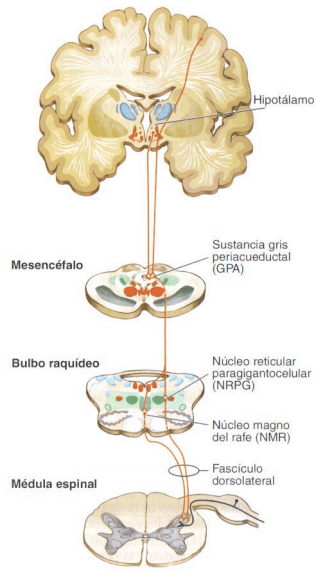
La vía neural más importante está constituida por los axones que alcanzan la sustancia gris periacueductal (GPA) donde establecen sinapsis con las neuronas del núcleo reticular paragigantocelular (NRPG) y del núcleo magno del rafe (NMR). Los axones que parten de estos núcleos se dirigen hacia la médula espinal formando parte del fascículo dorsolateral y alcanzando el asta dorsal a diferentes niveles medulares, donde modulan la transmisión de la información nociceptiva.
La existencia en el SNC de una serie de sustancias químicas que actúan como analgésicos endógenos, los péptidos opioides que se unen a receptores específico ampliamente distribuidos por todo el SNC, los receptores opioides.
Existen diferentes tipos de péptidos opioides que se localizan en zonas implicadas en el procesamiento o modulación del dolor, entre ellas la GPA. Las neuronas de la GPA reciben a su vez proyecciones descendentes que proceden de distintas zonas encefálicas como el hipotálamo, diferentes núcleos del tálamo, la amígdala y la corteza frontal.
Estas proyecciones proporcionan un sustrato neurobiológico que podría explicar la supresión del dolor que se da en determinadas situaciones. Los sistemas de analgesia son activados para permitir la realización de conductas de importancia biológica.
Diversos factores cognitivos y emocionales ponen en marcha los circuitos neurales que pueden suprimir la percepción del dolor. El dolor puede ser inhibido en situaciones de estrés: analgesia inducida por estrés.
8. Funciones Biológicas de los Sentidos
La función más evidente de los sistemas sensoriales es la de proporcionarnos información sobre el medio ambiente en que vivimos, lo que nos permite interaccionar con él. Otras funciones tienen el objetivo final de favorecer la adaptación del individuo a su entorno, lo que garantiza en definitiva su supervivencia.
Fase cefálica: consiste en el conjunto de reflejos digestivos (secreción de saliva, de insulina, secreciones gástricas…) que se ponen en marcha al ser estimulados los receptores sensoriales, sobre todo del olfato y del gusto, junto con otros receptores sensoriales de la boca y del encéfalo.
Los factores cefálicos correlacionan especialmente con la calidad de la comida: a mejor comida mayor amplitud de los reflejos cefálicos. Cuando se eliminan las sensaciones cefálicas asociadas a la ingestión mediante lesión nerviosa o por bloqueo farmacológico se eliminan también las diferencias en la respuesta cefálica.
Por otro lado, el gusto sirve bastante bien a la función de seleccionar adecuadamente la dieta y existen 2 hechos que lo corroboran: lo que se conoce como hambres específicas y el fenómeno comportamental llamado aversión gustativa.
La aversión gustativa consiste en que un animal aprende a evitar una comida determinada si en alguna ocasión su ingestión vino seguida al cabo de minutos o incluso horas, de sensaciones desagradables (mareos, vómitos). La aversión gustativa se basa en la asociación entre el sabor de la comida ingerida, sea el que sea, con el malestar posterior.
La selección natural es la responsable de que la gratificación por ingerir alimentos que contienen elementos esenciales de los que el organismo está falto sea mayor que cuando se ingieren estos mismos alimentos en situaciones normales de dieta equilibrada habitual.
El gusto es esencial para el control de lo que ingerimos, pero también el olfato desempeña una función relevante en la adquisición de la aversión gustativa. El olfato juega un papel protagonista en el despliegue de la conducta sexual de casi todos los mamíferos, incluido el hombre.
Las sustancias odoríferas o feromonas son mensajeros químicos que interactúan con receptores específicos desencadenando determinados procesos fisiológicos o promoviendo conductas específicas. Las feromonas afectan directamente a la secreción de hormonas hipotalámicas e hipofisarias.
Las proyecciones límbicas del sistema olfatorio humano pueden explicar el efecto que las feromonas tienen en nuestra especie.