

A los diferentes sistemas usados tanto para actuar sobre el medio externo como para controlar nuestro estado interno se les denomina sistemas efectores.
Uno de estos sistemas efectores es el sistema motor somático o simplemente sistema motor. Este sistema motor está organizado de modo jerárquico y está constituido por los músculos y los circuitos neurales que ordenan los movimientos, de forma que es el Sistema Nervioso Central quien coordina todas las respuestas que constituyen nuestro repertorio conductual.
Otros sistemas efectores son el sistema nervioso autónomo (SN autónomo) y el sistema endocrino.
Las respuestas de los sistemas efectores están bajo el control último del SNC que actúa de forma muy estrecha y en coordinación con el sistema endocrino, cuando nos referimos a estos 2 sistemas de forma conjunta se denomina sistema neuroendocrino.
Las diferentes respuestas emitidas por el sistema motor, el SN autónomo y el sistema endocrino están mediadas por diferentes tipos de órganos que se conocen con el nombre de órganos efectores.
Tipos de efectores
Hay diversos tipos de órganos efectores encargados de configurar diversos tipos de respuestas, son principalmente las glándulas y los músculos, que se corresponden respectivamente con los 2 tipos de acciones efectoras: la secreción glandular y la contracción muscular. Las glándulas son los órganos formados por células secretoras que cuentan con una maquinaria especializada, el aparato de Golgi, encargado de almacenar, concentrar y empaquetar sustancias específicas en vesículas denominadas gránulos de secreción.
Estos gránulos de secreción permanecen en el citoplasma de la célula hasta que ésta es estimulada, liberando sus secreciones fuera de la glándula.
Existen 2 tipos de glándulas:
– Glándulas endocrinas: sintetizan hormonas que son liberadas a la circulación sanguínea para actuar sobre células y órganos diana situados a distancia en el interior del organismo.
– Glándulas exocrinas: segregan sus productos en conductos especiales que los transportan a órganos adyacentes o al medio externo.
La mayoría de estas glándulas están controladas de alguna forma por el SN, principalmente por el SN autónomo.
En las glándulas que están bajo control nervioso, la estimulación de la célula ganglionar es producida por el neurotransmisor liberado desde la neurona eferente, que generalmente provoca su despolarización.
Los mecanismos son similares a los implicados en la liberación del neurotransmisor desde la célula nerviosa.
Figura 12.1 Esquemas representativos de las semejanzas en los mecanismos que conducen a la liberación de sustancias desde las células glandulares bajo control nervioso (izquierda) y desde las células nerviosas (a la derecha).

Los músculos también pueden ser de diferentes tipos, principalmente, estriados y lisos.
Los músculos estriados están controlados por neuronas localizadas en el SNC y su estimulación produce la contracción muscular, base de los movimientos. Se denominan también músculos esqueléticos, ya que todos los músculos que se fijan al esqueleto por medio de tendones son de este tipo.

El movimiento equivalente a la apertura de la navaja se denomina extensión y a los músculos cuya contracción es responsable de este movimiento se les denomina músculos extensores.
El movimiento en la dirección que cierra la navaja se denomina flexión y los músculos responsables se denominan músculos flexores.
Los músculos que actúan juntos para mover la articulación en una dirección son músculos sinérgicos entre sí.
Dado que flexores y extensores mueven la articulación en direcciones opuestas, se dice que son músculos antagonistas entre sí.
Los músculos esqueléticos están formados por numerosas fibras que se extienden en toda su longitud, las fibras musculares1.
Cada fibra muscular está delimitada por una membrana celular excitable denominada sarcolema y está compuesta por miofibrillas pudiendo llegar a tener varios miles de miofibrillas.
Cada miofibrilla consta de una cadena de pequeñas unidades repetidas regularmente, denominadas sarcómeros, que constituyen las unidades contráctiles de la fibra y confieren a la miofibrilla su apariencia estriada (bandas claras y oscuras).
Los músculos lisos deben su denominación a que no presentan las estrías del músculo esquelético. Están compuestos por fibras mucho más pequeñas y de menor longitud, no presentan miofibrillas ni la organización del músculo esquelético.
El músculo cardíaco tiene características que lo sitúan entre el liso y el estriado. Está compuesto por miofibrillas similares a las del músculo esquelético, diferenciándose fundamentalmente en su disposición.
Los músculos estriados son los efectores del sistema motor, mientras que los músculos lisos y el músculo cardíaco son efectores del SN autónomo. En el caso de las glándulas, se trata de órganos efectores tanto del SN autónomo como del sistema endocrino.
Aproximación General a los Sistemas Motores
Las posturas y movimientos se alternan constantemente en el comportamiento y ambos se encuentran bajo control del SNC, que planifica, coordina y ejecuta el control motor de las estructuras periféricas (músculos esqueléticos) que intervienen en su mantenimiento o realización.
Gran parte del control motor depende de diversas estructuras del encéfalo que canalizan sus señales a través de vías descendentes que confluyen finalmente en las motoneuronas espinales y troncoencefálicas. Estas neuronas motoras inervan los músculos esqueléticos y su activación desencadena la contracción muscular, base de los distintos movimientos.
Hay diferentes tipos de movimientos:
- Los movimientos reflejos: son las unidades elementales del comportamiento motor. Se ejecutan de modo continuado para mantener las condiciones posturales necesarias que van a permitir el desarrollo de conductas más complejas. Son respuestas relativamente simples, rápidas, estereotipadas e involuntarias, cuyo inicio puede ser desencadenado por estímulos sensoriales y que una vez que se han disparado no pueden ser modificadas hasta que terminan. En el reflejo más sencillo la neurona sensorial establece sinapsis directamente sobre la neurona motora, sin intervención de otros grupos neuronales del SNC.
- Los movimientos voluntarios: son os que realizamos con un propósito, requieren una planificación previa y son los más complejos. Son movimientos en gran medida aprendidos cuya ejecución mejora con la práctica, de tal forma que pueden realizarse de forma casi inconsciente.
Los movimientos rítmicos: no son típicamente reflejos ni típicamente voluntarios. Se realizan de forma automática y no requieren nuestra atención, a no ser que aparezcan perturbaciones inesperadas, en cuyo caso, podemos iniciarlos o terminarlos voluntariamente, aunque una vez iniciados, la secuencia de movimientos repetitivos se ejecuta de forma automática.
Una de las características fundamentales de los sistemas motores es que muestran una organización jerárquica, lo que significa que existe una gradación de funciones entre sus distintos componentes, de forma que las órdenes fluyen desde los niveles superiores a los inferiores.
El nivel inferior en el organigrama de control motor está constituido por las motoneuronas de la médula espinal y del tronco del encéfalo, que tienen autonomía para realizar actos motores estereotipados y automáticos, como los movimientos reflejos, sin contar con los niveles superiores, aunque también reciben órdenes motoras desde ellos.
El nivel superior de la jerarquía motora lo desempeña la corteza cerebral, en concreto, la corteza motora y diversas áreas de asociación.
Desde la corteza motora se originan vías motoras descendientes de la corteza cerebral al tronco del encéfalo y a la médula espinal.
El nivel intermedio de esta jerarquía motora está constituido por diversos núcleos del tronco del encéfalo, donde también se originan parte de los sistemas descendentes a la médula espinal.
Hay 2 sistemas moduladores: Los ganglios basales y el cerebelo que no envían órdenes directas a las motoneuronas sino que intervienen en el control motor modulando la actividad de los sistemas descendentes.

Esta organización jerárquica por la que las órdenes descienden desde los niveles superiores a los inferiores pasando por niveles intermedios se denomina organización en serie. Sin embargo, las vías motoras descendentes no sólo están organizadas en serie, sino que existe una organización directa en paralelo, de forma que existen vías que van directamente de la corteza cerebral a la medula espinal.
Esta organización en paralelo aporta mayor capacidad de procesamiento y de adaptación en el control motor.

Las Neuronas motoras y los receptores sensoriales de los músculos
Gran parte del control motor es llevado a cabo por diversas estructuras encefálicas, una parte de este control se realiza en circuitos locales localizados en la médula espinal y en el tronco del encéfalo.
Para que un movimiento pueda ser ejecutado de forma correcta, el SNC debe recibir información que le permita conocer la posición de las extremidades y el estado en que se encuentran los músculos antes de iniciar los movimientos y durante su ejecución.
Los receptores que realizan esta función son los propioceptores y están situados en los músculos y las articulaciones.
Las Motoneuronas y la contracción muscular
Las motoneuronas o neuronas motoras establecen sinapsis con las fibras musculares. A través de estas sinapsis, la información procedente de diferentes zonas del SNC se transmite de forma unificada a los músculos, transformándose en una respuesta muscular.
Las motoneuronas encargadas de esta función son las motoneuronas alfa que se localizan en las astas anteriores o ventrales de la medula espinal y en los núcleos motores del tronco del encéfalo.
La sinapsis que se establece entre el botón terminal de una neurona motora y la membrana de una fibra muscular constituye la unión neuromuscular.
Cada motoneurona, su axón y las fibras musculares que inerva constituyen una unidad motora, que es la unidad básica de funcionamiento que subyace a cualquier conducta motora.
Figura 12.6 Unidad motora formada por una motoneurona alfa localizada en la médula espinal, el axón eferente que llega hasta el músculo y las fibras musculares con las que éste establece sinapsis.

Los botones sinápticos localizados en las ramificaciones del axón penetran hasta una región especializada de la membrana de la fibra muscular denominada placa terminal (o placa motora).
Figura 12.7 La unión neuromuscular. Cuando el axón de la neurona motora alcanza el músculo pierde su envoltorio de mielina y se ramifica en un complejo de terminales nerviosos que se invaginan en la fibra muscular.

El neurotransmisor liberado desde las zonas activas de los botones sinápticos es la acetilcolina (ACh) y los receptores que se localizan en la membrana de la fibra muscular son receptores nicotínicos.
Cuando un potencial de acción llega a los botones terminales del axón de la motoneurona, se da la liberación de la ACh. La unión de este neurotransmisor a sus receptores en la fibra muscular abre los canales para sodio y potasio.
Figura 12.8 La ACh liberada desde el axón de la motoneurona alfa produce un potencial de placa en la fibra muscular como consecuencia de la apertura de los canales de sodio y potasio activados por neurotransmisor (1). Esta despolarización abre los canales de sodio activados por voltaje (2). Con la entrada de suficiente sodio, se da la despolarización necesaria para disparar un potencial de acción y producir la contracción de la fibra muscular (3).

La entrada y salida de estos iones produce una despolarización de la membrana postsináptica que recibe el nombre de potencial de placa terminal.
Los potenciales de placa terminal se diferencian de los potenciales excitatorios postsinàpticos producidos en las sinapsis entre neuronas, en que los primeros son lo suficientemente amplios para genera un potencial de acción que provoca la contracción de la fibra muscular.

Las sinapsis de la unión neuromuscular son siempre excitatorias, lo que implica que un músculo sólo puede estar relajado cuando se suprime la actividad de la neurona motora.
Desde el punto de vista funcional, existen distintos tipos de fibras musculares:
– Fibras de contracción rápida: responden de una forma veloz y enérgica pero se fatigan con rapidez.
– Fibras de contracción lenta: mantienen una contracción más duradera, alcanzan su máximo nivel más lentamente y presentan mayor resistencia a la fatiga.
Cada músculo está compuesto por fibras de contracción rápida y fibras de contracción lenta.
En cuanto a la fuerza con la que se produce la contracción muscular, la fuerza de contracción depende del número de unidades motoras que se activan y de la frecuencia de los potenciales de acción disparados por la motoneurona.
Cuanto mayor es el número de unidades motoras activadas y mayor es la frecuencia de descarga de potenciales de acción, mayor será la fuerza de contracción.
En cuanto a la precisión con que controlamos nuestros movimientos depende de la cantidad de axones motores que alcanzan los diferentes grupos musculares y del número de fibras musculares que cada uno inerva.
El control será más preciso cuanto menor sea el número de fibras musculares inervadas por un axón.
Receptores sensoriales de los músculos. La propiocepción
El organismo dispone de receptores especializados que nos informan de las deformaciones mecánicas que se producen en el interior de nuestro cuerpo y que nos permiten conocer cuándo y cuánto se estiran y contraen nuestros músculos.
Estos son los propioceptores, (receptores de sí mismo), mientras que el término propiocepción hace referencia a los mecanismos sensoriales que nos informan de los movimientos y de la posición de nuestro cuerpo.
Esta información llega al SNC desde los propioceptores localizados en los músculos, los tendones y las articulaciones. Entre estos receptores se encuentran los mecanorreceptores de las articulaciones y los receptores sensoriales que se localizan en los músculos: los husos musculares y los órganos tendinosos de Golgi.
Husos Musculares
Se sitúan en paralelo entre las fibras que constituyen el músculo esquelético.
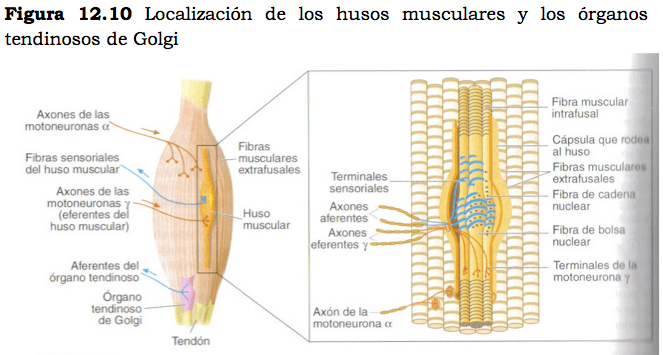
La densidad de husos varía en función de las características funcionales de cada músculo, de forma que los músculos más grandes y que producen movimientos menos precisos tienen pocos husos y aquellos que intervienen en los movimientos más finos cuentan con una mayor densidad.
En los husos musculares se pueden distinguir 3 componentes fundamentales: las fibras intrafusales y los terminales sensoriales y motores.
Cada huso muscular está compuesto de 2 a 14 fibras musculares especializadas denominadas fibras intrafusales (dentro del huso) rodeadas por una cápsula de tejido conectivo.
Alrededor del huso muscular se localizan las fibras musculares, también llamadas fibras extrafusales que constituyen los elementos contráctiles del músculo.
La información captada por el huso muscular sobre el estado de las fibras musculares que lo rodean es transmitida al SNC por axones sensoriales mielinizados que establecen sinapsis con la porción central de las fibras intrafusales.
Cuando se da el estiramiento o extensión de un músculo, los husos musculares, también se estiran.
El estiramiento del huso alarga la zona central de las fibras intrafusales, produciendo la activación de canales iónicos sensibles al estiramiento, la despolarización de los terminales sensoriales y el disparo de potenciales de acción en los axones aferentes que transmiten la información al SNC.
Cuando se da el estiramiento, los husos envían información al SNC pero cuando se da la contracción de las fibras musculares que hace que se acorten al igual que las fibras intrafusales y como consecuencia, los husos dejan de enviar información al SNC.
Para que esto no ocurra el SNC dispone de mecanismos de control centrífugo que le permiten regular la información que recibe enviando axones eferentes a los mismos órganos sensoriales de los que proceden las aferencias.
Las motoneuronas gamma (o neuronas fusimotoras), están localizadas en el asta ventral unto a las motoneuronas alfa. A través de ellas el SNC modula la información sensorial que le llega desde los músculos, de forma que los husos musculares están inervados por axones eferentes mielinizados pertenecientes a estas motoneuronas.
Durante la realización de los movimientos, la estimulación de las motoneuronas alfa se compensa con la activación conjunta de las motoneuronas gamma. Esta coactivación es fundamental para enviar al SNC información acerca de si el movimiento se está dando correctamente y para que el SNC pueda compensar los posibles errores.
La activación de las motoneuronas gamma asegura que las aferencias del huso continúen transmitiendo información, debido a que mantienen el estiramiento de las fibras intrafusales y en consecuencia regulan la sensibilidad de los receptores que proporcionan información sobre el estado de los músculos.
Órganos Tendinosos de Golgi
Son receptores encapsulados localizados en la unión del músculo y el tendón, que se sitúan en serie con las fibras musculares extrafusales a las que se unen mediante fibras de colágeno trenzadas.
Estos propioceptores están inervados por axones sensoriales mielinizados que se ramifican entre las fibras de colágeno y que son algo más finos que aquellos que inervan los husos musculares.

La diferente disposición de los husos musculares (en paralelo) y de los órganos tendinosos de Golgi (en serie) respecto a las fibras musculares extrafusales determinan respuestas muy distintas al estiramiento y a la contracción del músculo.
La actividad en las aferencias procedentes de los husos musculares transmite al SNC información sobre la longitud del músculo (estiramiento) mientras que la actividad en las aferencias procedentes de los órganos tendinosos de Golgi transmite información sobre la tensión muscular (contracción).
Los Reflejos medulares
Los reflejos son las unidades elementales del comportamiento motor. Se trata de respuestas simples, rápidas y estereotipadas en cuya ejecución participan 5 componentes:
- – Un receptor sensorial
- – Una vía aferente hacia el SNC
- – Una o más sinapsis en el SNC
- – Una vía eferente
- – Un efector
Componentes básicos de un reflejo

La complejidad de los reflejos varía en función de los grupos neuronales que intervienen, siendo el reflejo más sencillo aquel en el que la neurona sensorial establece sinapsis directamente con la neurona motora, sin intervención de otros grupos neuronales del SNC.
Los reflejos elicitados por circuitos neurales tanto del tronco del encéfalo como de la médula espinal son los reflejos medulares.
Entre los reflejos medulares el más sencillo es el reflejo de extensión o miotático que es elicitado por la extensión de un músculo, de forma que ésta provoca la contracción refleja del mismo, como si se resistiera al estiramiento.
Figura 12.15 Utilidad del reflejo de extensión: el estiramiento del músculo que se da al aplicar una carga, como cuando sostenemos una jarra llena, provoca su propia contracción, como si se resistiera al estiramiento, para evitar la caída de esa carga.


Los axones aferentes también establecen sinapsis con interneuronas inhibitorias que disminuyen la actividad de las motoneuronas alfa que inervan los músculos antagonistas.
Al disminuir la estimulación de estos músculos, también decrece la tensión y se da el estiramiento.
Mecanismo de la inhibición recíproca: un grupo de músculos es excitado al mismo tiempo que sus antagonistas son inhibidos.
El circuito neural que constituye el reflejo de extensión tiene una única sinapsis, la que establecen las fibras aferentes del huso muscular con las motoneuronas alfa, al reflejo de extensión también se le denomina reflejo monosináptico.
Los reflejos de extensión permiten mantener un cierto grado de contracción de la musculatura extensora, el tono muscular, que es la fuerza por la que el músculo se resiste a la extensión producida por la gravedad.
Reflejos Polisinápticos
El reflejo de extensión es el único reflejo medula en el que interviene una sola sinapsis, todos los demás son reflejos polisinápticos ya que están controlados por circuitos donde una varias interneuronas se intercalan entre la neurona sensorial y la motora.
Estos reflejos varían en cuanto a su complejidad. Algunos son relativamente sencillos como el reflejo de flexión y el reflejo miotático inverso.
El reflejo de flexión se desencadena en su grado máximo cuando se aplica a una extremidad un estímulo nocivo, lo que provoca la contracción de todos los músculos flexores de la extremidad para retirarla del estímulo, se denomina también reflejo de retirada.
Puede implicar a diferentes músculos en función de la intensidad del estímulo.
Figura 12.17 Circuito neural del reflejo de flexión o de retirada y del reflejo de extensión cruzado.

La estimulación de los receptores cutáneos produce la contracción de os músculos flexores y la inhibición de los músculos extensores ipsilaterales, lo que permite retirar la extremidad.
En algunas ocasiones la retirada del miembro donde se ha dado la estimulación nociva produce la respuesta opuesta en los músculos del miembro contralateral, de forma que se contraen los músculos extensores y se inhiben los músculos flexores para mantener la extremidad apoyada. A este reflejo se le denomina reflejo de extensión cruzado.
Mientras el miembro dañado se aparta del peligro, el otro miembro mantiene el equilibrio o participa en la locomoción.
El reflejo miotático inverso, en el cual los procesos involucrados son opuestos a los que se dan en el reflejo de extensión o miotático. Su función es enlentecer la contracción muscular según va aumentando su magnitud y disminuirla si ésta es tan elevada que existe riesgo de que se lesionen los tendones por un estiramiento excesivo.
Cuando esto ocurre, los órganos tendinosos de Golgi envían información a la médula espinal para activar el reflejo de forma que se dé la inhibición de los músculos flexores, lo que evita la contracción y la estimulación de los extensores para permitir el estiramiento del brazo.
Figura 12.18 Circuito neural del reflejo miotático inverso. Este circuito de retroalimentación negativa regula el grado de la contracción muscular.

Areas corticales que intervienen en el control motor
Sólo una parte del control motor se lleva a cabo en estos circuitos locales responsables de los reflejos.
Sin embargo, la ejecución de la mayoría de nuestros movimientos está bajo control de diversas estructuras del encéfalo, que constituyen los niveles superiores e intermedios de la jerarquía motora y cuyas señales confluyen finalmente en las motoneuronas espinales y troncoencefálicas.
El nivel superior de esta jerarquía está constituido por la corteza cerebral (corteza motora y diversas áreas de asociación). En la corteza cerebral se localizan 2 componentes esenciales de los sistemas motores:
– las áreas de asociación de la corteza parietal posterior y prefrontal dorsolateral.
– las áreas motoras, constituidas por las áreas motoras secundarias o áreas premotoras (zona lateral) y el área motora primaria.
Áreas de asociación de la corteza cerebral
Las señales procesadas en la corteza parietal posterior se transmiten a la corteza de asociación prefrontal dorsolateral y a las áreas motoras de la corteza.
Figura 12.19 Flujo de señales aferentes y eferentes entre las diversas áreas corticales implicadas en el control motor

La corteza de asociación parietal posterior parece intervenir en los movimientos dirigidos a un blanco aportando las claves sensoriales necesarias para su realización y las señales motivaciones relacionadas con el estado del individuo, enviando esta información a la corteza de asociación prefrontal dorsolateral y a la corteza motora.
La corteza de asociación prefrontal dorsolateral desempeña un papel fundamental en la planificación de nuestro comportamiento en función de la experiencia.
La información que recibe desde la corteza de asociación parietal posterior es almacenada en esta área cortical para aportar una representación mental de los estímulos a los que el sujeto tiene que responder.
Esta corteza prefrontal dorsolateral selecciona los aspectos más relevantes la estrategia más adecuada para ejecutar con éxito el movimiento. Además, toma la decisión de iniciar los movimientos.
Áreas motoras de la corteza cerebral
Las áreas motoras de la corteza cerebral son un componente esencial y exclusivo de los sistemas motores y desde ellas parten las vías descendentes que envían las órdenes a los niveles inferiores de la jerarquía motora.
Estas áreas se localizan en el lóbulo frontal, anteriores a la cisura central.
El área o corteza motora primaria se localiza en la circunvolución precentral, desde la cisura lateral hasta la superficie medial del hemisferio cerebral.
Anterior a ella, se localizan las áreas premotoras o corteza motora secundaria, constituidas por la corteza premotora en la superficie lateral del hemisferio y el área motora suplementaria, en la parte superior y medial del mismo. (Figura 12.19 C).
Figura 12.20 Organización somatotópica de la corteza motora primaria y homúnculo motor.

La corteza motora primaria, al igual que otras áreas corticales esta organizada en columnas que controlan la acción de músculos individuales.
Las zonas que controlan cada parte del cuerpo en las 3 áreas de la corteza motora de cada hemisferio están interconectadas por fibras de asociación.
– Función de las áreas premotoras o corteza motora secundaria
La función primordial de la corteza premotora es participar en la preparación de los movimientos, en la planificación o programación motora, especialmente de los movimientos desencadenados por estímulos externos.
El área premotora se activa cuando se realiza un movimiento guiado por un estímulo externo pero si no existe esta estimulación externa, la corteza premotora no se activa aunque el sujeto realiza el movimiento.
Las neuronas del área motora suplementaria también se activan antes de que se ejecute una tarea motora y también requiere una mayor estimulación que el área motora primaria para evocar movimientos.
El área motora suplementaria también interviene en la programación motora y además en la coordinación de movimientos complejos (coordinación bimanual)
Función del área motora primaria
El área motora primaria se activa durante la realización de los movimientos y sobre ella convergen las proyecciones de las áreas premotoras por lo que parece que estas áreas la preparan para su ejecución. Además recibe importantes proyecciones desde otras áreas.
El área motora primaria participa en el inicio o disparo del movimiento.
Las neuronas individuales de la corteza motora primaria codifican el grado de fuerza ejercida por los músculos, la fuerza de la contracción muscular, mediante un código temporal, por la frecuencia de disparo de potenciales de acción.
Algunas de estas neuronas corticales parece que codifican también la velocidad de la fuerza que se está aplicando.
La dirección en la que se han de realizar los movimientos no parece depender tanto de la activación de neuronas individuales, sino de la acción conjunta de amplias poblaciones de neuronas que responden de forma desigual a diferentes direcciones del movimiento.
Se considera que la corteza motora primaria elabora las órdenes motoras de cuándo y cómo se han de mover los músculos para enviarlas a través de los sistemas motores descendentes (junto a las señales procedentes de las áreas premotoras) a los niveles inferiores de la jerarquía motora, las motoneuronas que inervan los músculos, para que se produzca la contracción muscular y la ejecución de los movimientos.
Sistemas Motores Descendentes
Los sistemas motores descendentes están formados por las vías que descienden desde la corteza motora y desde el tronco del encéfalo hasta confluir sobre las motoneuronas que ocupan el nivel inferior de la jerarquía motora y son la vía final común que controla la contracción de los músculos.
Esta organización en paralelo de vía descendentes indirectas (en serie) y directas aporta una mayor capacidad de procesamiento y de adaptación en el control motor.
Sistemas Descendentes de la Corteza Motora
Los axones que se originan en la corteza motora y se dirigen al tronco del encéfalo y a la médula espinal forman varios tractos que descienden organizados de forma somatotópica e influyen sobre las motoneuronas troncoencefálicas y espinales que inervan los músculos representados en sus zonas de origen.
Figura 12.24 Zonas de origen de los tractos motores en el área motora primaria y organización somatotópica en su trayectoria descendentes

En las zonas de la corteza motora donde están representadas la cabeza y la cara se origina el tracto corticobulbar. A través de este tracto la corteza motora controla los músculos de la cabeza que intervienen en movimientos voluntarios.
Hay vías que descienden en posición lateral y otras que lo hacen en posición medial:
- – Las vías laterales son el tracto corticoespinal lateral (vía directa) y las fibras corticorrubrales (vía indirecta)
- Las vías mediales son el tracto corticoespinal ventral (vía directa) y las fibras corticorreticulares (vía indirecta)
Figura 12.25 Vías descendentes de la corteza motora primaria y de las áreas premotoras

Los tractos corticoespinales son los más largos del SNC y a través de ellos la corteza motora influye de forma directa sobre las motoneuronas de la médula espinal.
Se originan en las 3 áreas motoras, pero contiene tantos axones procedentes del área motora primaria como la suma de los que proceden de las áreas premotoras.
El tracto corticoespinal lateral se origina en las zonas de la corteza motora en las que están representadas las extremidades (antebrazos, parte inferior de las piernas, manos, pies y dedos). Este tracto cruza la línea media en las pirámides bulbares. Al ser un tracto cruzado, el tracto corticoespinal lateral controla los músculos de las extremidades contralaterales.
El tracto corticoespinal ventral se origina en las zonas de la corteza motora en las que están representados el cuello, el tronco y las zonas proximales de las extremidades (cadera, hombro o el brazo). Este tracto no cruza la línea media en las pirámides.
La corteza motora influye también sobre la médula espinal a través de 2 vías indirectas que terminan sobre diversos núcleos del tronco del encéfalo que envían proyecciones descendentes a la médula espinal. Estas vías descendentes de la corteza motora terminan en el núcleo rojo (fibras corticorrubrales) y en la formación reticular (fibras corticorreticulares).
Se originan en las mismas zonas de representación cortical que los tractos corticoespinales, por lo que las que terminan en el núcleo rojo proceden de las zonas de representación de las extremidades al igual que el tracto corticoespinal lateral, pues ambas son vías laterales. Mientras que las que terminan en la formación reticular se originan en las zonas de representación del cuello, del tronco y de las zonas proximales de las extremidades, al igual que el tracto corticoespinal ventral, pues ambas son vías mediales.
Sistemas descendentes del tronco del encéfalo
En el tronco del encéfalo se originan diversos tractos formados por axones que se dirigen a la médula espinal y que al igual que los procedentes de la corteza motora, se organizan en vías que descienden en posición lateral o medial.
La vía lateral se origina en el núcleo rojo, tracto rubroespinal. Al igual que el tracto corticoespinal lateral, este tracto sigue una trayectoria cruzada.
Figura 12.26 Vía lateral del tronco del encéfalo: tracto rubroespinal

Las vías mediales están constituidas por diversos tractos originados en diferentes núcleos del tronco del encéfalo (como la formación reticular, los núcleos vestibulares y el colículo superior). Estas vías descienden en posición ventral al igual que el tracto corticoespinal ventral, que es también otra vía medial
Figura 12.27 Vías mediales del tronco del encéfalo: tratos vestibuloespinal medial y lateral.

Funciones de los sistemas descendentes
Los sistemas motores descendentes de la corteza motora y del tronco del encéfalo a la médula espinal llevan a cabo diferentes funciones según se trate de las vías laterales o mediales.
Estas funciones son:
- – Promover los movimientos independientes de las extremidades para la ejecución de movimientos voluntarios, llevada a cabo por las vías laterales.
- – Controlarla postura.
- – Dirigir la locomoción.
Estas 2 últimas funciones son responsabilidad de las vías mediales.
Las funciones concretas de las vías laterales están constituidas por una vía directa, el tracto corticoespinal lateral y una vía indirecta a través del
tronco del encéfalo (fibras corticorrubrales y tracto rubroespinal).
El tracto corticoespinal lateral es la vía fundamental para la realización de movimientos voluntarios, finos y precisos de las extremidades, sobre todo de las manos.
Una de sus funciones primordiales es controlar los movimientos fraccionados (discretos) o independientes de los dedos.
El tracto corticoespinal lateral interviene también en otros aspectos del control motor, como es la preparación de los músculos de las extremidades para iniciar movimientos voluntarios.
La otra vía lateral (indirecta) descendente de la corteza motora al núcleo rojo (fibras corticorrubrales) y desde éste a la médula espinal (tracto rubroespinal) parece participar también en el control voluntario de las extremidades contralaterales, en concreto, de los movimientos independientes de los hombros y de las extremidades, fundamentalmente del codo y de la mano, pero no de los dedos.
Las funciones de las vías mediales está constituidas por el tracto corticoespinal ventral (vía directa) y diversos tractos que van de la corteza cerebral al tronco del encéfalo y desde éste a la médula espinal (indirecta).
El tracto corticoespinal ventral es la vía directa por la que la corteza motora interviene en el control de los músculos del cuello, del tronco y de los músculos proximales de las extremidades, y colabora con la otra vía medial (indirecta) en el control de la postura y de la locomoción.
Las vías mediales del tronco del encéfalo desempeñan un papel fundamental en estas funciones, ya que distribuyen las señales de control muchos núcleos motores de la médula espinal relacionados con el mantenimiento del equilibrio y de la postura.
Cada uno de los tractos que forman las vías mediales del tronco del encéfalo realiza su propia contribución al mantenimiento de la postura, aunque se considera que los núcleos reticulares del puente son la región facilitadora por excelencia de los reflejos espinales antigravitatorios para el mantenimiento de la postura erguida.
Además de las vías mediales del tronco del encéfalo controlan los ajustes posturales anticipatorios que requiere cualquier movimiento voluntario.
Las vías mediales del tronco del encéfalo también participan en el control de los movimientos que permiten desplazarse de un sitio a otro, denominados en conjunto locomoción y que requieren la coordinación de contracciones alternas y rítmicas de los músculos extensores y flexores.
Los patrones motores rítmicos implicados en la locomoción se generan en la médula espinal, en unos circuitos o redes neurales denominados generadores de acción central o generadores centrales de patrones. En el tronco del encéfalo residen los centros que activan estos generadores centrales de patrones rítmicos de la locomoción, concretamente en la región locomotora subtalámica y en la región locomotora mesencefálica.
Por tanto, la médula espinal y los sistemas descendentes del tronco del encéfalo aportan patrones elementales de control motor que permiten que los niveles superiores puedan centrar su actividad en controlar aspectos más complejos.
Resumen: Las vías laterales son las únicas que cruzan al lado opuestos del cuerpo para controlar los músculos de las partes más laterales de las extremidades. Por el contrario, las vías mediales no cruzan al lado opuesto por lo que discurren medialmente para controlar los músculos del tronco y de las partes más proximales de las extremidades y precisamente por su disposición y función, son las más indicadas para participar en el control de la postura y en la locomoción.
Sistemas moduladores: El cerebelo y los ganglios basales
Además de los 3 niveles de la jerarquía motora, hay otros 2 componentes de los sistemas motores que desempeñan una importante función en el control motor: el cerebelo y los ganglios basales.
Estos no envían vía directamente a la médula espinal, sino que ejercen su influencia principalmente a través de conexiones con los componentes donde se originan los sistemas motores descendentes, por lo que son considerados centros moduladores del control motor.
El Cerebelo
Esta estructura compara las órdenes motoras descendentes que se están emitiendo, con la información acerca de los movimientos que se están realizando.
En función del resultado, actúa a través de diferentes vías sobre el tronco del encéfalo y la corteza motora para aportar precisión a los movimientos y corregir los posibles errores.
El cerebelo puede intervenir, ya sea modulando la actividad de las vías descendentes o bien, la actividad de la corteza premotora, modificando los programas motores.
Las 3 divisiones funcionales del cerebelo actúan en paralelo para controlar diferentes aspectos del control motor mediante su influencia sobre los sistemas motores descendentes.
Figura 12.31 Influencias de las divisiones funcionales del cerebelo sobre las vías motoras descendentes

El VESTIBULOCEREBELO envía señales correctoras a los núcleos vestibulares para modificar la postura y restablecer el equilibrio.
Las lesiones en esta zona producen inestabilidad tanto al mantenerse quieto de pie como durante la ejecución de movimientos.
El ESPINOCEREBELO genera señales que corrigen las disonancias entre la acción que se pretende realizar y la que se está llevando a cabo en los músculos.
Cada uno de los componentes de esta zona influye sobre distintas vías descendentes modificando el tono muscular de los músculos laterales de las extremidades e interviniendo en el control de la postura y en la locomoción.
Las lesiones en esta zona provocan un característico modo de andar vacilante o titubeante.
El CEREBROCEREBELO es la unidad funcional que modula principalmente los sistemas motores descendentes de la corteza cerebral para la planificación de nuevos movimientos y su ejecución suave y precisa. Su influencia es clave para la coordinación de los movimientos voluntarios. Participa en la preparación de los movimientos, sobre todo de los movimientos en los que intervienen múltiples articulaciones y de los movimientos fraccionados de los dedos, en asociación con la corteza premotora y envía a la corteza motora primaria órdenes importantes para disparar el inicio de los mismos.
Las lesiones en esta zona provocan demoras en el inicio y en la terminación de los movimientos, desórdenes en la coordinación temporal de aquellos en los que intervienen múltiples articulaciones… Su función fundamental es por tanto la programación de secuencias coordinadas de movimientos y su coordinación temporal.
Los Ganglios Basales
Los ganglios basales (cuerpo estriado, núcleo subtalámico y sustancia negra) forman un sistema funcional de gran importancia para el control motor.
La alteración de los ganglios basales produce trastornos motores que implican la aparición de movimientos involuntarios. 2 de estos trastornos son la corea o enfermedad de Huntington y la enfermedad de Parkinson.
Al igual que ocurre con el cerebelo, los ganglios basales no forman parte de los sistemas motores descendentes sino que su intervención en el control motor se realiza por bucles de retroalimentación entre los diversos componentes y, en conjunto, a través del tálamo, con la corteza de asociación prefrontal y con la corteza motora sobre todo con el área motora suplementaria.
Los ganglios basales intervienen fundamentalmente en la planificación y en la fase de inicio de los movimientos.
El neoestriado es el principal centro receptor de las señales que llegan al cuerpo estriado, mientras que el globo pálido y la sustancia negra reticulada son los centros efectores del sistema de los ganglios basales, de modo que dentro del sistema las señales viajan desde el neoestriado hacia los 2 centros efectores.
Figura 12.32 2 circuitos de retroalimentación de los ganglios basales a la corteza cerebral.

En el neoestriado existe una gran diversidad de circuitos neuroquímicos a través de los que éste recibe señales excitadoras e inhibidoras.
Figura 12.33 Modelo de funcionamiento normal de los ganglios basales

Los trastornos motores hipercinéticos como la corea de Huntington, el balismo que es un trastorno que se caracteriza por un movimiento de lanzamiento violento e involuntario de las extremidades o los tics, caracterizados por las contracciones estereotipadas, repetitivas e involuntarias de determinados músculos, parecen depender de una disminución en la actividad del núcleo subtalámico (NST)
Los trastornos hipocinéticos, de los que la enfermedad de Parkinson es el prototipo, se producen por la degeneración de la sustancia negra. La degeneración de gran parte de las neuronas dopaminérgicas de la sustancia negra y de sus proyecciones al neoestriado produce cambios en la actividad de los circuitos de este sistema, dando como resultado un debilitamiento de las señales que llegan a las áreas mototas de la corteza cerebral, al producirse una reducción de las señales excitatorias procedentes del tálamo.
Figura 12.34 Modelo de funcionamiento de los ganglios basales en la enfermedad de Parkinson

El Sistema Nervioso Autónomo
El sistema nervioso autónomo (SNA) forma parte del SNP y constituye otro de los sistemas efectores.
Su función es regular la actividad de los órganos internos para ajustar su funcionamiento frente a las demandas del medio.
Se necesitan mecanismos fisiológicos coordinados que permitan mantener un estado de equilibrio interno u homeostasis, por lo que los mecanismos neurales esenciales para mantener esta estabilidad se localizan en el hipotálamo.
Esta estructura del SN integra la información que recibe de muchas regiones del encéfalo y genera una respuesta unificada a través de los 2 sistemas que están bajo su control: el SNA y el sistema endocrino.
Las respuestas del SNA para mantener la homeostasis están mediadas por diferentes órganos efectores: el músculo cardíaco, los músculos lisos situados en diferentes órganos y las glándulas.
El SNA incluye fibras aferentes que llevan información sensorial al SNC sobre el estado de los órganos internos para modular la actividad autónoma.
Los cuerpos celulares de los que se originan estos axones aferentes se localizan en los ganglios de las raíces dorsales de la médula espinal y en los ganglios de los nervios craneales.
El SNA es considerado principalmente un sistema eferente, un sistema de respuesta. Las fibras eferentes del SNA ejercen el control de la musculatura lisa, del músculo cardíaco y de las glándulas.
El SNA no funciona de forma autónoma, también se le denomina involuntario o vegetativo. Generalmente las funciones controladas por el SNA suelen llevarse a cabo de una forma automática, y de una manera extraordinariamente coordinada.
Figura 12.35 Organización del SNA. Presenta 2 divisiones: el SN simpático y el SN parasimpático

Organización anatómica del SNA
El SNA controla el funcionamiento de los órganos internos mediante reflejos con diferente grado de complejidad. Los hay relativamente simples y en ellos intervienen la médula espinal y el tronco del encéfalo, mientras que otros requieren un control más complejo dependiente de niveles superiores del SNC.
Algunos núcleos del tronco del encéfalo contribuyen a regular el SNA: el núcleo del tracto solitario es uno de ellos. Este núcleo del bulbo raquídeo conectado con el hipotálamo, recibe la información sensorial procedente de las vísceras y controla el funcionamiento del SNA a través de un conjunto de circuitos reflejos.
Otras funciones del SNA requieren de la regulación de centro superiores del SNC, siendo el hipotálamo la estructura encefálica que desempeña el papel más relevante en este control.
Este centro de control integra las aferencias que recibe desde diferentes regiones del encéfalo y la información visceral procedente de la médula espinal y del tronco del encéfalo para generar un patrón coherente de respuestas autónomas.
En la inervación autónoma intervienen 2 neuronas que unen el SNC con los órganos de la periferia, la neurona preganglionar y la neurona postganglionar.
El cuerpo celular de la neurona preganglionar se localiza en el SNC; mientras que el de la neurona postganglionar se encuentra en el ganglio autónomo.
Figura 12.36 Esquema en el que se comparan las conexiones que se establecen entre el SNC y los órganos efectores del sistema motor somático y del SNA

Las neuronas que emiten las órdenes motoras a los músculos estriados se localizan en el SNC (en la médula espinal o en el tronco del encéfalo) mientras que las neuronas que inervan a los órganos efectores del SNA se localizan fuera del SNC, en los ganglios autónomos.
El SNA tiene 2 divisiones principales: el SN simpático y el SN parasimpático, existiendo diferencias fundamentales en su organización anatómica y funcional:
– El lugar del SNC donde se localizan las neuronas preganglionares:
En el SN simpático los axones preganglionares se localizan en las astas laterales de la médula espinal y en el SN parasimpático se localizan en núcleos del tronco del encéfalo y en algunos segmentos sacros intermedios de la médula espinal.
– La localización de los ganglios periféricos:
En el SN simpático los axones preganglionares son cortos, porque establecen sinapsis con ganglios muy próximos al SNC y en el SN parasimpático los axones preganglionares son generalmente largos situándose cerca de los órganos que inerva o en ellos.
– Los neurotransmisores:
Los liberados por los axones postganglionares se dan sobre los órganos diana. Los axones postganglionares del SN simpático liberan principalmente noradrenalina que se une a diferentes subtipos de receptores adrenérgicos.
Los axones postganglionares del SN parasimpático liberan acetilcolina que se une a los receptores muscarínicos presentes en las membranas de las células efectoras.
Hay similitudes también entre el SN simpático y parasimpático ya que en ambos casos las neuronas preganglionares liberan acetilcolina que se une a receptores nicotínicos situados en todos los ganglios autónomos.
Una tercera división del SNA es el sistema nervioso entérico que se encuentra situada en su totalidad en la periferia y su actuación muestra una gran autonomía respecto al hipotálamo y al resto del SNC. Este sistema neural proporciona inervación a los órganos del sistema digestivo.
Está formado por una red de neuronas interconectadas que constituyen 2 plexos cuyos somas se localizan en múltiples ganglios. Recibe aferencias del SN simpático y parasimpático y también indirectamente está modulado por el SNC.